")
![]()
Mizer is an R package to run dynamic multi-species size-spectrum models of fish communities. The package has been developed to model marine ecosystems that are subject to fishing. However, it may also be appropriate for other aquatic ecosystems. By providing a framework for multi-species fisheries modelling as an R package, mizer enhances the accessibility, usability, and reproducibility of models, and thus aims to facilitate collaboration and innovation.
The package contains functions that allow you to set up an ecosystem model and then project it through time under different fishing strategies. Methods are included to explore the results, including plots and calculations of community indicators such as the slope of the size spectrum. Size-based models can be complicated, so mizer contains many default options that you can however change when needed.

Mizer has been supporting research in marine ecology and fisheries science since 2014 (see publications). Mizer is still under active development. Version 2 has increased the user-friendliness and the flexibility of the framework. Version 3 has introduced diffusion, arising from stochastic growth, and improved the numerics. Contributions from the user community are very welcome. There is a sister package called mizerExperimental where user contributions can be checked out and receive feedback from the community. A growing number of other extension packages add new biology such as temperature dependence, starvation mortality, seasonal dynamics, and more — see the list of extension packages. Example mizer models can be contributed to mizerExamples. Follow us on twitter and read our blog to stay up-to-date with new developments. Does your project or publication use mizer? If so, we would love to know.
Recent work on mizer was funded by the European Commission Horizon 2020 Research and Innovation Programme under Grant Agreement No 634495 for the project MINOUW and the Australian Research Council Discovery Project Rewiring Marine Food Webs.
Installation
The package is on CRAN and therefore available from R’s built-in package manager.
# Install latest released version from CRAN
install.packages("mizer")
# Alternatively, install the development version from GitHub
pak::pak("sizespectrum/mizer")Example
The following code loads the mizer package, loads some information about species in the North Sea that comes as an example with the package, sets up the parameters for the mizer model, and runs a simulation for 10 years.
library(mizer)
params <- newMultispeciesParams(NS_species_params, NS_interaction)
sim <- project(params, t_max = 10, effort = 0)The results of the simulation can then be analysed, for example via plots:
plot(sim)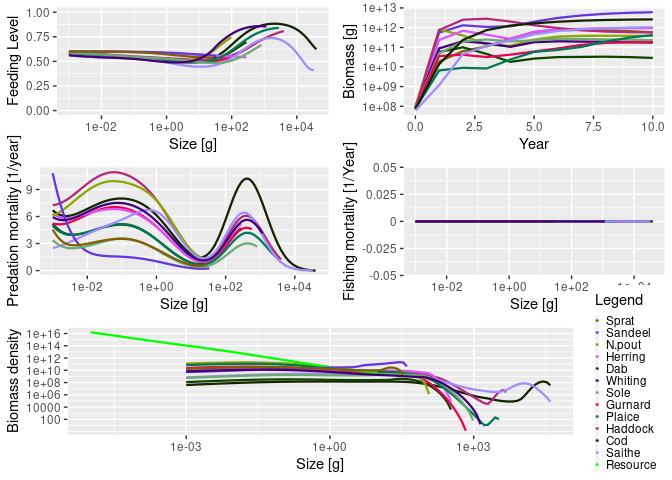
See the accompanying Get started page for more details on how the package works, including detailed examples.

Dynamic multi-species size-spectrum model
Size-based multi-species models are important for fisheries science because they provide a more realistic and accurate representation of the dynamics of fish populations and the ecosystems in which they live. In contrast to traditional single-species models, which consider a single fish stock as an isolated unit, size-based multi-species models account for the fact that fish populations are part of a larger ecosystem and interact with other species through predation, competition, and other ecological processes.
One of the key advantages of size-based multi-species models is that they provide a more comprehensive understanding of the impacts of fishing on fish populations and ecosystems. By considering the size distribution of different fish species, these models can capture the effects of fishing on both target and non-target species, and on different life stages of a species. This is particularly important for species that are caught as bycatch or that are indirectly affected by fishing through changes in their food web.
Another advantage of size-based multi-species models is that they can be used to investigate the effects of environmental changes and other perturbations on fish populations and ecosystems. For example, these models can be used to explore the impacts of climate change on the distribution and abundance of fish populations, or the effects of habitat loss or pollution on fish communities. Because mizer is a mechanistic model, it can deduce the complex population-level changes that we are interested in from the simpler changes in the physiological rates and feeding interactions of individual fish species. You can find examples of this in the list of publications.
Overall, size-based multi-species models provide a more comprehensive and realistic framework for understanding the dynamics of fish populations and ecosystems, and for developing effective fisheries management strategies that account for the complex interactions among species and their environment.

A mizer model captures the interactions between species. The growth rates of fish are determined by the availability of prey and the death rates are influenced by the abundance of predators, as well as fishing. The model starts with the individual-level physiological rates for each species, as well as the predation preferences, and deduces the population-level dynamics from these. Thus quantities like fish diets and fisheries yields emerge dynamically and can be projected into the future.
Because a mizer model tracks the size of individuals as they grow up over several orders of magnitude from their egg size to their maximum size, it correctly tracks the ontogenetic diet shifts. An individual typically moves through several trophic levels during its life time. This is often not correctly captured in other multi-species models.
A mizer model can be set up with only a small amount of information because it uses allometric scaling relations and size-based feeding rules to choose sensible defaults for unknown parameters.
Setting up a new multi-species mizer model is a two-step process, similar to what may be familiar from Ecopath with Ecosim: First one calibrates the model to describe a steady state that is in agreement with current observations (as in Ecopath), then one chooses the additional parameters that determine the dynamics away from the steady state (as in Ecosim). This model can then be used to investigate future effects of changes in fishing policy or of environmental stressors.
A strong theoretical basis
One big advantage of a mizer model is that it is based on a strong mathematical foundation. This allows a degree of a priori understanding of the behaviour of the model that is absent in many other multi-species models. This theoretical foundation is well presented in the book “Fish Ecology, Evolution, and Exploitation” by Ken Andersen.
It is interesting to think of the marine ecosystem as a transport system that moves biomass from the size of primary producers (mostly unicellular plankton) up to the sizes of fish that humans like to consume. Each fish that grows up from egg size to maturity by eating smaller individuals is like a car on this biomass highway. The yield of our fisheries depend on this traffic flowing smoothly and without traffic jams.
An analogy with road traffic may be helpful:

In road traffic, if traffic density gets too high in a section of the highway, drivers slow down, which leads to a pile-up producing even higher traffic density, leading to further slow-down in a potentially vicious cycle known as a traffic jam. Traffic management that ignores how the traffic density affects traffic speed fails. Luckily our mathematical understanding of transport equations has made practical contributions to managing traffic in ways that produce smoother traffic flow and hence higher throughput.
Mizer implements the transport equations for marine ecosystems. The potential for traffic jams is the same: if for example there is a high density of predators of a particular size, which all have preference for prey of a particular smaller size, then due to competition for that prey the growth of those predators slows down, leading to a pile-up which leads to further depletion of prey, leading to further slow-down, in a potentially vicious cycle.
Luckily, the natural ecosystem has evolved to facilitate very smooth traffic on this biomass highway, with resultant high productivity. This state is characterised by an approximate power-law shape of the biomass size spectrum. The purpose of mizer is to allow us to understand how various stressors, like fishing or climate change, affect the size spectrum and hence the flow of biomass and the productivity and resilience of the marine ecosystem. Mizer allows us to investigate how size-based fisheries management strategies can be used to keep the ecosystem close to its natural productive state.