The simplest version of the size spectrum model is the community model. In the community model, individuals are only characterized by their size and are represented by a single group representing an across-species aggregate. Reproduction is not considered; the reproduction rate \(R\) is set to be constant. The resource spectrum only extends to the start of the community spectrum. Metabolism is turned off.
In this section we describe how a community model can be set up and projected through time. We then use a community model to illustrate the idea of a trophic cascade. Due to the relative simplicity of this type of model, it is useful for gently introducing some of the concepts behind the mizer package. Consequently, this section should hopefully serve as an introduction to using mizer.
Setting up a community model
The first stage in implementing a model using mizer is
to create an object of class MizerParams. This class
contains the model parameters including the life-history parameters of
the species in the model, the fishing selectivity functions and the
parameters of the resource spectrum.
To avoid having to make a MizerParams object directly,
the newCommunityParams() wrapper function, has been
provided that conveniently creates a MizerParams object
specifically for a community model. The documentation for the function
can be seen by clicking on the function name anywhere it appears on this
page: newCommunityParams().
As can be seen in the help page, the function can take many arguments. We can ignore most of these for the moment as they almost all come with default values.
The arguments that you should pay attention to are: z0
(the external mortality rate), alpha (the assimilation
efficiency of the community), f0 (the average feeding level
of the community, which is used to calculate \(\gamma\)) and h (the
coefficient of the maximum intake rate).
Although default values for these parameters are provided, you are
encouraged to explore how changing the values affects the simulated
community. For example, the default value of z0 is \(0.1\). Increasing this value effectively
‘shortens’ the length of the community spectrum.
The newCommunityParams() function is called by passing
in the arguments by name. Any parameter that is not passed in is set to
the default value. For example, the following line sets up the
parameters with z0 = 0.05, f0 = 0.5. All other
parameters will have their default value:
params <- newCommunityParams(z0 = 0.05, f0 = 0.5)Calling the function creates and returns an object of type
MizerParams. We can check this using the
class() function.
class(params)## [1] "MizerParams"
## attr(,"package")
## [1] "mizer"To work with mizer you do not have to worry about how this object is realised internally. All you need to know is that it contains all the information needed to run model simulations and that mizer provides you with functions to access, use and modify this information.
A very brief description of the model contained in the MizerParams
object can be seen by calling the summary() method on
it:
summary(params)## An object of class "MizerParams"
## mizer version: 3.2.1
## Created: 2026-07-31 10:36:45
## Modified: 2026-07-31 10:36:45
## Consumer size spectrum:
## minimum size: 0.001
## maximum size: 1e+06
## no. size bins: 100
## Resource size spectrum:
## minimum size: 8.11131e-11
## maximum size: 0.000811131
## no. size bins: 78 (178 size bins in total)
## Species details:
## An object of class "species_params" containing parameters for 1 species:
## species w_inf w_mat w_min f0 beta sigma
## Community 1e+06 250000 0.001 0.5 100 2
##
## Fishing gear details:
## Gear Effort Target species
## ----------------------------------
## Community 0.00 CommunityIn the summary you can see that the size range of the community
spectrum has been set from \(0.001\) to
\(10^{6}\) and this is divided into
\(100\) size bins. Similar information
is available for the resource spectrum. Additionally, the community is
made up of only one species, called Community, which has an
maximum size of \(10^{6}\) and a
preferred predator prey mass ratio of \(100\). The w_mat parameter has
been set to NA as it is not used when running a community
model. These values have all been set by default using the
newCommunityParams() function.
Running the community model
The newCommunityParams() function has given us a
MizerParams object that contains all the information mizer
needs about the model community. We can use this to perform a simulation
and project the community through time. In the mizer
package, projections are performed using the project()
function. You can see the help page for project() for more
details and it is described fully in the section on running a
simulation. We will ignore the details for the moment and just use
project() to run some simple projections. The arguments for
project() that we need to be concerned with are
effort, which determines the fishing effort (and therefore
fishing mortality) through time, and t_max, which is the
number of years we want to project into the future. Initial population
abundances have been set automatically (by the
get_initial_n() function). It is possible to set your own
initial abundances but we will not do this here.
To run a projection for 50 years, with no fishing effort (i.e. we want to model an unexploited community) we run:
sim <- project(params, t_max = 50, effort = 0)The resulting object, sim, is of type
MizerSim.
class(sim)## [1] "MizerSim"
## attr(,"package")
## [1] "mizer"This class holds the results of the simulation, including the community and resource abundances at size through time, as well as the original model parameters. It is explained in detail in the section on running a simulation.
After running the projection, it is possible to explore the results using a range of plots and analyses. These are described fully in the section on exploring the simulation results.
To quickly look at the results of the projection you can call the
plot() method. This plots the feeding level, predation
mortality, fishing mortality and abundance by size in the last time step
of the simulation, and the total biomass through time. Each of the plots
can be shown individually if desired.
plot(sim)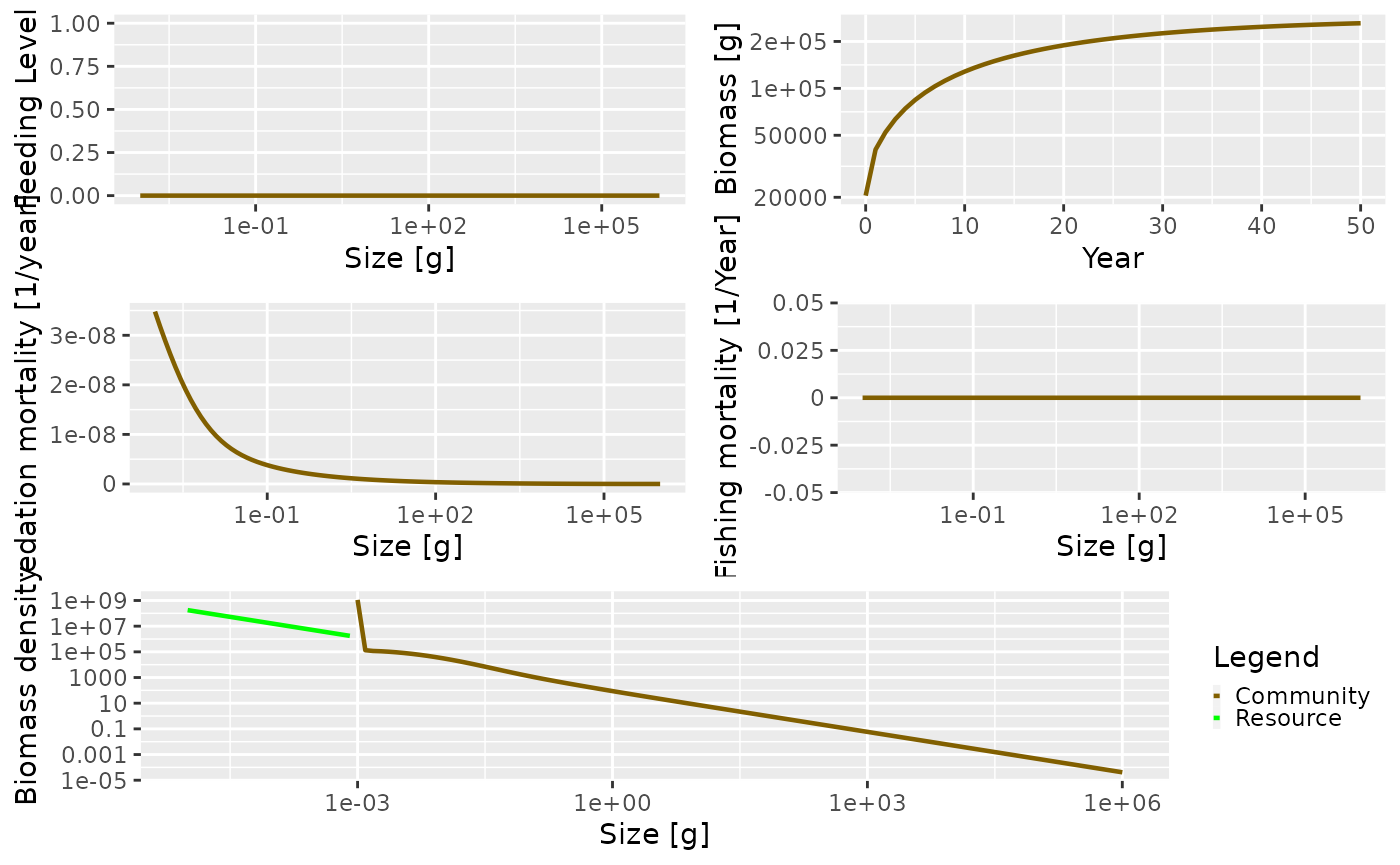
In the above plot there are several things going on that are worth
talking about. Looking at the total biomass of the community against
time, you can see that the biomass quickly reaches a stable equilibrium.
The other panels show what is happening at the last time step in the
simulation, which in this case is when the community is at equilibrium.
Fishing mortality is 0 because we set the effort argument
to 0 when running the simulation. The predation mortality rate is
clearly a function of size, with the smallest sizes experiencing the
highest levels of predation. The feeding level describes how satiated an
individual is, with 0 being unfed, and 1 being full satiated. The
feeding level at size will be strongly affected by the values of the
f0 and alpha arguments passed to the
newCommunityParams() function.
The resource and community spectra are shown in the bottom panel of
the plot (the plotted resource spectrum has been truncated to make for a
better plot, but really extends all the way back to \(8.11\times 10^{-11}\) g). You can see that
the community spectrum forms a continuum with the resource spectrum.
This is strongly affected by the level of fixed reproduction rate (the
reproduction argument passed to
newCommunityParams())
Note the hump in the biomass at the largest end of the community
spectrum. This is because the size spectrum model can be broadly
described as ‘big things eating little things’. Given this, what is
eating the very biggest things? Without fishing pressure, the mortality
of the largest individuals is only from the external mortality
(determined by the z0 argument) and the mortality from
predation is almost 0. This is difficult to see in the plot due to the
predation mortality being so high for the smaller individuals.
We can see this more clearly by extracting the predation mortality
information from the MizerSim object, sim,
that we created above. This is easily done by using the
getPredMort() function (see the help page for more
details). There are several functions that can be used for extracting
information from a MizerSim object,
e.g. getFeedingLevel() and getFMort(). For
more information see the
section on exploring the simulation results. Here we just call
getPredMort() using the sim object:
pred_mort <- getPredMort(sim)The resulting pred_mort object is an array that contains
the predation mortality at time by species by size. Here we only have
one species so the species dimension is dropped, leaving us with a two
dimensional array of time by size. We projected the model for \(50\) time steps but the length of the time
dimension is \(51\) as the initial
population is also included as a time step. We can get the index of the
final time with
idxFinalT(sim)## [1] 51To pull out the predation mortality at size in the final time step we use:
pred_mort_final <- pred_mort[idxFinalT(sim), ]If you plot this predation mortality on a log-log scale you can see how the predation mortality declines to almost zero for the largest sizes.
plot(x = w(params), y = pred_mort_final, log = "xy", type = "l",
xlab = "Size [g]", ylab = "Predation mortality [1/year]")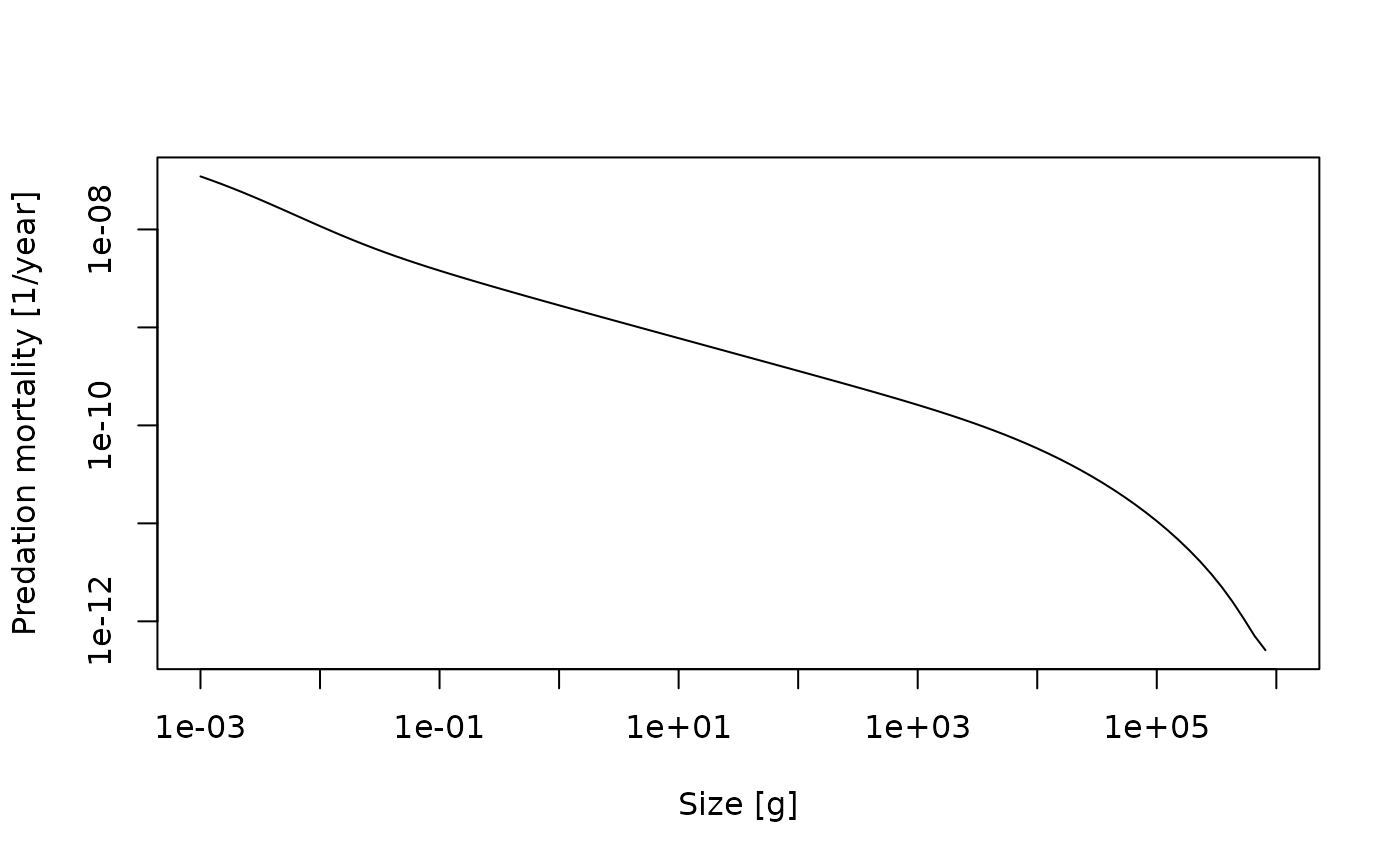
Note how we used w(sim) to get the vector of sizes to
plot along the x-axis and how we specified that we wanted to have
logarithmic axes.
In the long run it is worthwhile to use the ggplot2 package for
creating plots, so we show also how you would create the above graph
with ggplot(). For more detail see the section on using ggplot2 and plotly with mizer.
library(ggplot2)
sd <- data.frame(x = w(params), y = pred_mort_final)
ggplot(sd, aes(x = x, y = y)) +
geom_line() +
scale_x_log10() + scale_y_log10() +
xlab("Size [g]") + ylab("Predation mortality [1/year]")## Warning in scale_y_log10(): log-10 transformation introduced
## infinite values.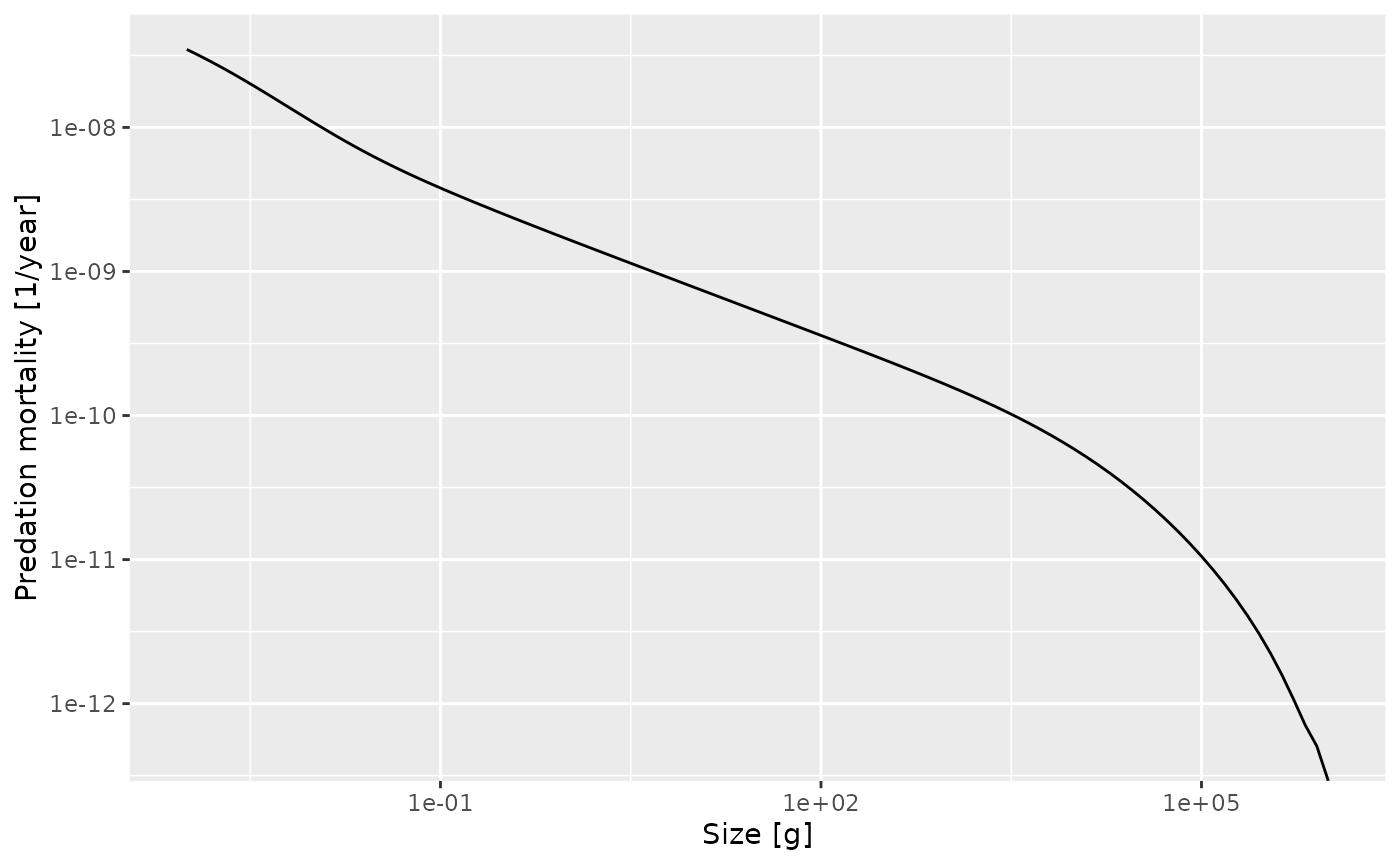
Example of a trophic cascade with the community model
It is possible to use the community model to simulate a trophic cascade. To do this we need to perform two simulations, one with fishing and one without.
This means we need to consider how fishing is handled in
mizer. The newCommunityParams() function
automatically sets the fishing selectivity to have a knife-edge shape,
with only individuals larger than 1 kg selected (the size at the
knife-edge can be changed by setting the knife_edge_size
argument). Although it is possible to change the selectivity function,
here we will use the default knife-edge selectivity. We set up the
parameter object with default parameters:
params_knife <- newCommunityParams()First we perform a simulation without fishing in the same way we did
above by setting the effort argument to 0:
sim0 <- project(params_knife, effort = 0, t_max = 50)Now we want to simulate again, this time with fishing. In the simulations, fishing mortality is calculated as the product of the fishing selectivity, effort and catchability (see the section on fishing gears for more details). By default catchability is set to 1. This means that a fishing effort of 1 will result in a fishing mortality of 1/year for fully selected sizes. Here we run a simulation with fishing effort set to 1 for the duration of the simulation:
sim1 <- project(params_knife, effort = 1, t_max = 50)You can compare the difference between these scenarios by using the
plot() method as before. Of particular interest is the
fishing mortality at size. The knife-edge selectivity at 1000 g can be
clearly seen and an effort of 1 has resulted in a fishing mortality of 1
for the fully selected sizes.
plot(sim1, power = 2)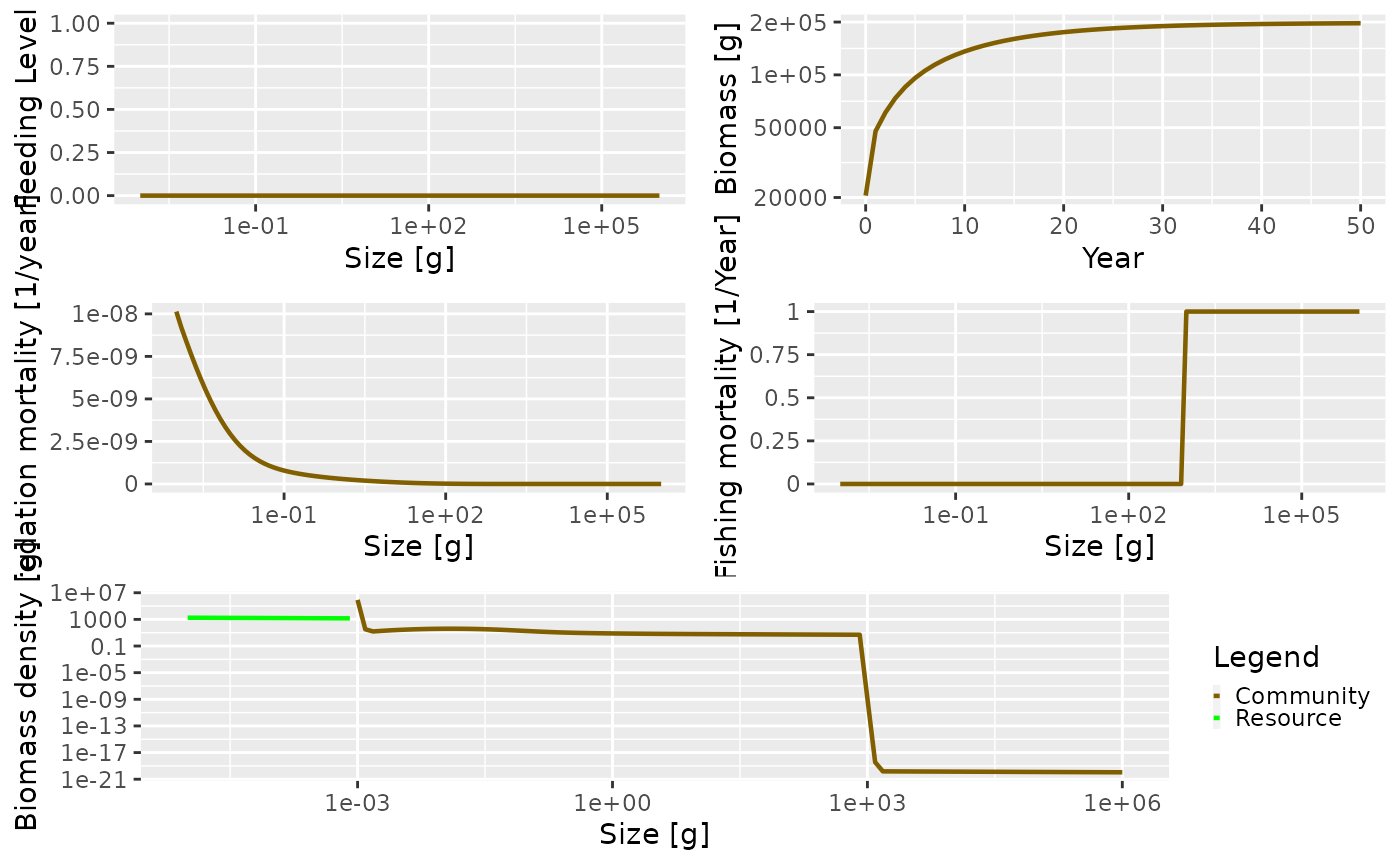
To explore the presence of a trophic cascade, we are interested in
looking at the relative change in abundance when the community is fished
compared to when it is not fished. To do this we need to get the
abundances at size from the simulation objects. This is done with the
N() function, which returns a three dimensional array with
dimensions time x species x size. Here we have 51 time steps (50 from
the simulation plus one which stores the initial population), 1 species
and 100 sizes:
## [1] 51 1 100We want the abundances in the final time step, and we can use these to calculate the relative abundances:
For convenience, to save you from having to determine the index of
the final time, mizer provides the function finalN(), so we
could have done
This can then be plotted using basic R plotting commands.
plot(x = w(params), y = relative_abundance, log = "x", type = "n",
xlab = "Size (g)", ylab = "Relative abundance")
lines(x = w(params), y = relative_abundance)
lines(x = c(min(w(params)), max(w(params))), y = c(1, 1), lty = 2)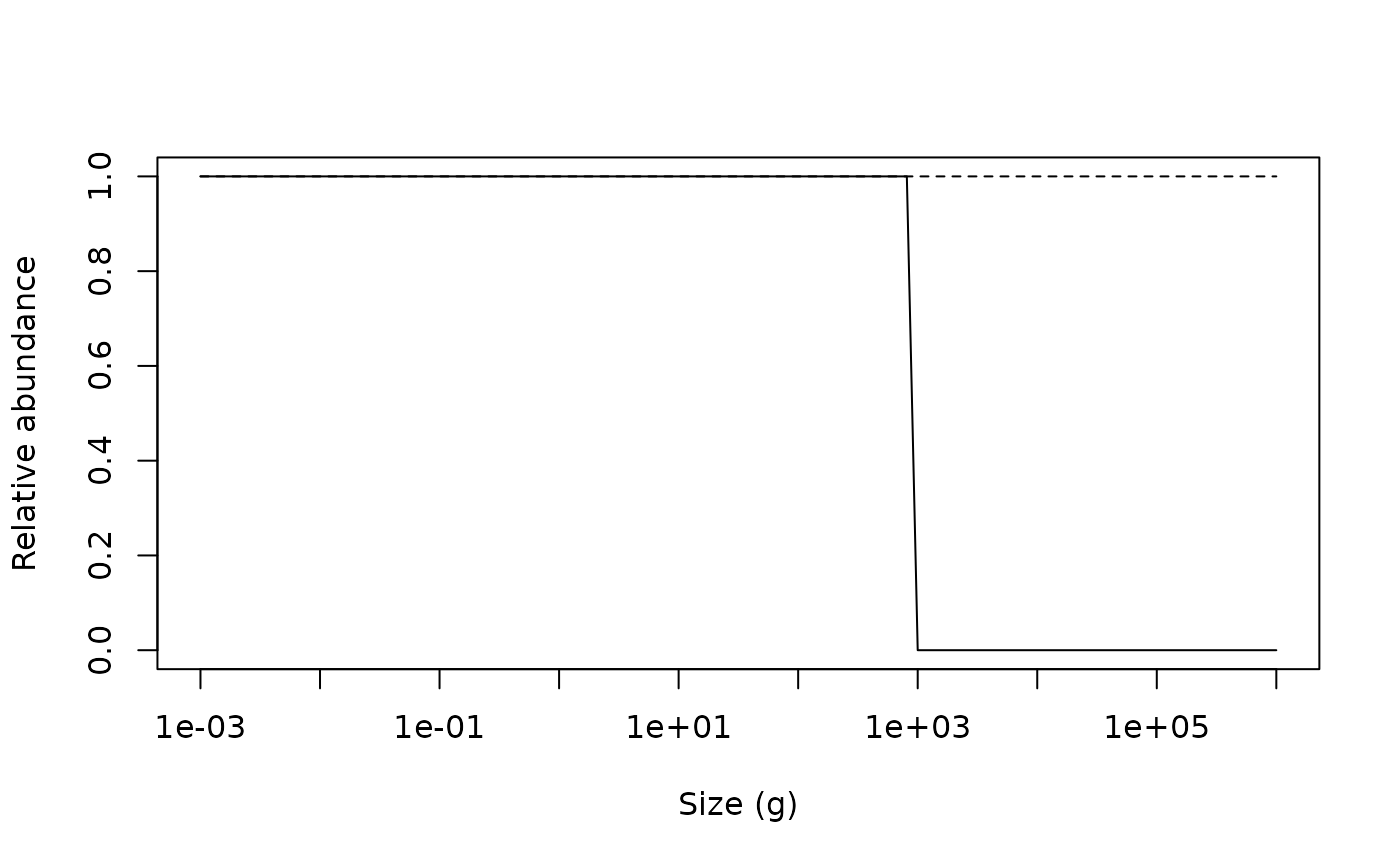
The impact of fishing on species larger than 1000g can be clearly seen. The fishing pressure lowers the abundance of large fish (the decrease in relative abundance at 1000 g). This then relieves the predation pressure on their smaller prey (the preferred predator-prey size ratio is given by the \(\beta\) parameter, which is set to 100 by default), leading to an increase in their abundance. This in turn increases the predation mortality on their smaller prey, which reduces their abundance and so on.
The impact of changing \(\sigma\)
As described above, the \(\sigma\) parameter determines the width of the predator prey size preference. Here we take a look at how changing the value of \(\sigma\) can affect the dynamics of the community.
In the examples above, \(\sigma\) is
set in the newCommunityParams() function by default to a
value of \(2\). We can see this by
looking at the sigma column of the species_params data
frame that is contained in the MizerParams object:
species_params(params)$sigma## Community
## 2When projected through time, the community abundances converge to a
stable equilibrium. What happens if we reduce the value of \(\sigma\), for example by setting it to 1.0?
We can do this by passing in the new value of \(\sigma\) into
newCommunityParams().
params_sigma1 <- newCommunityParams(sigma = 1)We want to project this new model through time using the
project() function. Here we project the new parameter
object for 50 time steps without fishing and save at intervals of 0.1
years (t_save = 0.1):
sim_sigma1 <- project(params_sigma1, effort = 0, t_max = 50,
dt = 0.01, t_save = 0.1)Note that we have introduced a new argument: \(dt\). This is the step size of the solver.
It does not have anything to do with the biology in the model. It only
affects the internal engine of project() that performs the
projection. As you can see in the underlying model equations in the model description section, the
model is formulated in continuous time. Therefore, to project it
forward, project() must solve the system of equations using
numerical methods. The quality of these methods is strongly affected by
\(dt\). The default value of \(dt\) is 0.1, which will be fine for most of
the projections we run in this document. Here it is necessary to reduce
the value to 0.01 to avoid introducing any artefacts into the projected
values. Decreasing \(dt\) increases the
time it takes to run a projection.
Let’s take a look at how the abundances change through time. We can
do this with the plotBiomass() function:
plotBiomass(sim_sigma1)
The plot above shows that abundances of the community no longer converge to a stable equilibrium and the dynamics appear to be chaotic. The ecological significance of the change in dynamics, and of the ability of simple community models to show chaotic behaviour, is still being debated. It can be argued that the size of the oscillations are too large to be ‘true’. Additionally, when a trait-based model is implemented, the magnitude of the oscillations are much smaller.
The next section is about the trait based model.